нуклеиновые основания представляют собой строительные блоки, из которых состоят длинные цепи молекул ДНК и РНК в их фосфорилированной нуклеотидной форме.
В ДНК, которая образует двойные цепи, похожие на веревки, 4 встречающихся нуклеиновых основания образуют твердые пары с соответствующим дополнительным основанием через водородные мостики. Нуклеотидные основания состоят либо из бициклического пуринового, либо из моноциклического пиримидинового скелета.
Что такое азотистые основания?
4 азотистых основания аденин, гуанин, цитозин и тимин, как строительные блоки длинных цепочек молекул с двойной спиралью ДНК, образуют постоянные пары аденин-тимин (A-T) и гуанин-цитозин (G-C).
Каждое из двух оснований, аденина и гуанина, состоит из модифицированного бициклического шести- и пятичленного кольца с основной пуриновой структурой, и поэтому их также называют пуриновыми основаниями. Основная структура двух других нуклеиновых оснований, цитозина и тимина, состоит из гетероциклического ароматического шестичленного кольца, которое соответствует модифицированному пиримидиновому скелету, поэтому их также называют пиримидиновыми основаниями. Поскольку РНК в основном представлена в виде одиночных цепей, изначально нет пар оснований. Это происходит только во время репликации через мРНК (информационная РНК).
Копия цепи РНК состоит из комплементарных азотистых оснований, аналогичных второй цепи ДНК. Единственная разница в том, что тимин в РНК заменен урацилом. Цепные молекулы ДНК и РНК не образуются в чистом виде из азотистых оснований, а вместо этого, в случае ДНК, соединяются с 5-сахарной дезоксирибозой с образованием соответствующего нуклеозида. В случае РНК группа сахара состоит из рибозы. Кроме того, нуклеозиды фосфорилируются фосфатным остатком с образованием так называемых нуклеотидов.
Пуриновые основания гипоксантин и ксантин, которые также встречаются в ДНК и РНК, соответствуют модифицированному тимину. Гипоксантин образуется из аденина путем замены аминогруппы (-NH3) гидроксильной группой (-OH), а ксантин образуется из гуанина. Оба азотистых основания не способствуют передаче генетической информации.
Функция, эффект и задачи
Одна из наиболее важных функций азотистых оснований, составляющих двойные цепи ДНК, - показать присутствие в заданном положении.
Последовательность азотистых оснований соответствует генетическому коду и определяет тип и последовательность аминокислот, из которых состоят белки. Это означает, что наиболее важная функция азотистых оснований как компонента ДНК состоит в пассивной, статической роли, то есть они не вмешиваются активно в метаболизм, и их биохимическая структура не изменяется под воздействием РНК-мессенджера (мРНК) в процессе считывания. Это частично объясняет долговечность ДНК.
Период полураспада митохондриальной ДНК (мтДНК), в течение которого распадается половина изначально существовавших связей между азотистыми основаниями, сильно зависит от условий окружающей среды и колеблется от примерно 520 лет в средних условиях с положительными температурами до 150 000 лет в условиях вечной мерзлоты. ,
В составе РНК азотистые основания играют несколько более активную роль. В принципе, когда клетки делятся, двойные цепи ДНК расщепляются и отделяются друг от друга, чтобы иметь возможность образовать дополнительную цепь, мРНК, которая, так сказать, образует рабочую копию генетического материала и служит основой для выбора и последовательности аминокислот, из которых собираются предполагаемые белки. Другое нуклеиновое основание, дигидроурацил, встречается только в так называемой транспортной РНК (тРНК), которая используется для транспортировки аминокислот во время синтеза белка.
Некоторые азотистые основания выполняют совершенно иную функцию как часть ферментов, которые активно каталитически активируют и контролируют определенные биохимические процессы. Аденин выполняет свою самую известную задачу в качестве нуклеотида в энергетическом балансе клеток. Аденин играет важную роль в качестве донора электронов, таких как аденозиндифосфат (АДФ) и аденозинтрифосфат (АТФ), а также компонент никотинамидадениндинуклеотида (НАД).
Образование, возникновение, свойства и оптимальные значения
В нефосфорилированной форме азотистые основания состоят исключительно из углерода, водорода и кислорода - веществ, которые распространены повсеместно и свободно доступны. Таким образом, организм способен сам синтезировать азотистые основания, но этот процесс является сложным и энергоемким.
Поэтому восстановление нуклеиновых кислот путем рециркуляции является предпочтительным, например Б. путем расщепления белков, содержащих определенные соединения, которые можно выделить и преобразовать в нуклеиновые кислоты с небольшими затратами энергии или даже с увеличением энергии. Нуклеиновые кислоты обычно встречаются в организме не в чистом виде, а в основном в виде нуклеозидов или дезоксинуклеозидов с присоединенной молекулой рибозы или дезоксирибозы. Как компонент ДНК и РНК, а также как компонент некоторых ферментов, нуклеиновые кислоты или их нуклеозиды также обратимо фосфорилируются одной-тремя фосфатными группами (PO4-).
Контрольного значения для оптимального количества азотистых оснований не существует. Дефицит или избыток азотистых оснований можно определить только косвенно, через определенные нарушения метаболизма.
Заболевания и расстройства
Опасности, нарушения и риски, связанные с азотистыми основаниями, представляют собой ошибки в количестве и последовательности цепей ДНК или РНК, которые приводят к изменению кодирования синтеза белка.
Если организм не может устранить неисправность с помощью своих механизмов восстановления, речь идет о синтезе биологически неактивных или пригодных для использования белков, что, в свою очередь, может привести к легким или серьезным метаболическим нарушениям. Это может, например, Присутствуют мутации гена B., которые могут вызвать симптоматические заболевания с самого начала через нарушения обмена веществ, которые могут быть неизлечимыми. Но даже в здоровом геноме ошибки копирования могут возникать во время репликации цепей ДНК и РНК, что влияет на метаболизм.
Известным нарушением метаболизма пуринового баланса является z. Б. вернулся к генетическому дефекту на x-хромосоме. Из-за генетического дефекта пуриновые основания, гипоксантин и гуанин, не могут быть переработаны, что в конечном итоге способствует образованию мочевых камней и подагре в суставах.

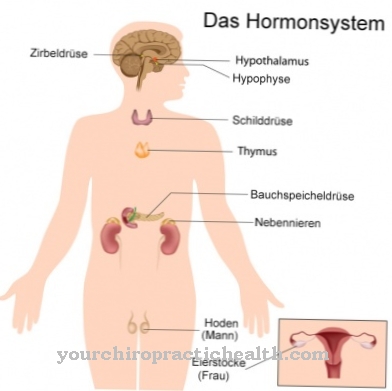



.jpg)
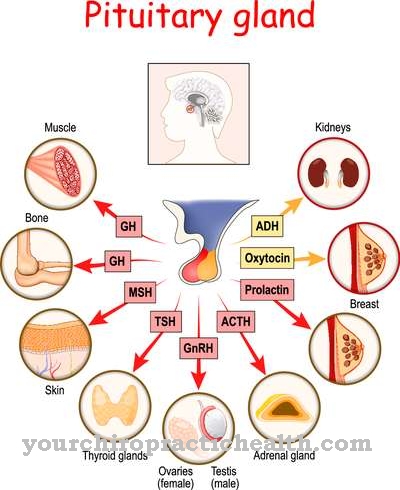




.jpg)