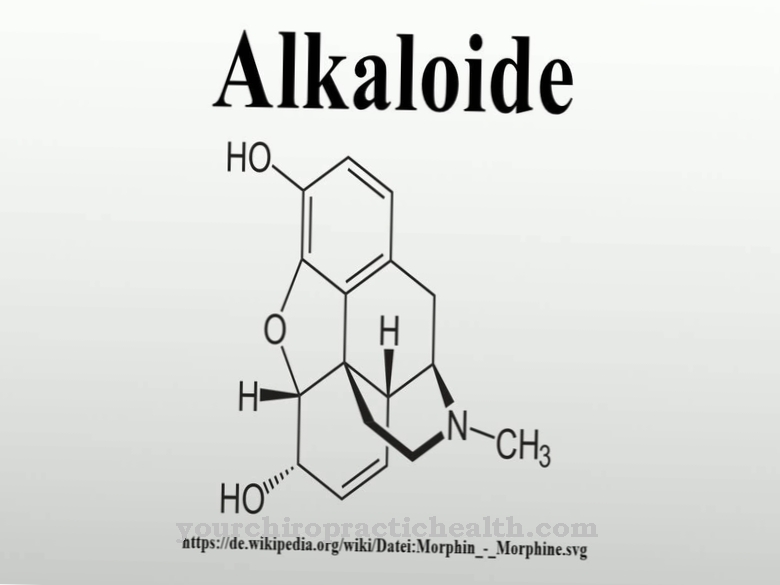
цитидин относится к нуклеозидам и состоит из цитозина нуклеинового основания и сахарной рибозы. Он образует пару оснований с гуанозином через водородные связи. Он также играет центральную роль в метаболизме пиримидина.
Что такое цитидин?
Цитидин - это нуклеозид, состоящий из цитозина и рибозы. Помимо аденина, гуанина и тимина, азотистое основание цитозина участвует в синтезе нуклеиновых кислот. Фосфорилирование цитидина дает цитидинмонофосфат (CMP), цитидиндифосфат (CDP) или цитидинтрифосфат (CTP).
Цитидинмонофосфат - это нуклеотид в РНК. Два пуриновых и два пиримидиновых основания участвуют в структуре нуклеиновых кислот, при этом тимин заменяется на урацил в РНК. Аденин и гуанин относятся к пуриновым основаниям, а тимин, цитозин и урацил относятся к пиримидиновым основаниям. Цитидиндезаминаза может дезаминировать цитидин до уридина. Уридин - это нуклеозид, состоящий из рибозы и урацила. Он также может фосфорилироваться до монофосфата уридина.
Монофосфат уридина также является важным нуклеотидом для РНК. Кроме того, CDP и CTP также являются активирующими группами для синтеза лецитина, цефалина и кардиолипина. Чистый цитидин присутствует в виде водорастворимого твердого вещества, которое разлагается при температуре от 201 до 220 градусов. Он может быть каталитически разложен до цитозина и рибозы под действием фермента пиримидиннуклеозидазы.
Функция, эффект и задачи
Цитидин играет центральную роль в метаболизме пиримидина. Пиримидин обеспечивает основную структуру пиримидиновых оснований цитозина, тимина и урацила, которые встречаются в нуклеиновых кислотах. Тимин в РНК заменен на урацил.
Урацил также производится дезаминированием цитидина цитидиндезаминазой. Химические превращения между тремя пиримидиновыми основаниями имеют центральное значение для процессов репарации в ДНК и эпигенетических изменений. В контексте эпигенетики различные свойства изменяются под влиянием окружающей среды. Однако генетический материал не меняется. Модификационные изменения организма вызываются разной экспрессией генов. Процессы дифференцировки клеток организма для образования различных клеточных линий и органов также представляют собой эпигенетический процесс: в зависимости от типа клеток активируются или деактивируются разные гены.
Это происходит за счет метилирования оснований цитидина в ДНК. Во время метилирования образуется метилцитозин, который путем дезаминирования может превращаться в тимин. Комплементарный гуанин азотистого основания в противоположной двойной цепи позволяет распознать ошибку и снова заменить тимин на цитозин. Однако гуанин также можно обменять на аденин, что приводит к точечной мутации. Если неметилированный цитозин дезаминирован, образуется урацил. Поскольку урацил не появляется в ДНК, он сразу замещается цитозином. Вместо цитозина немного увеличивается частота мутаций из-за метилирования.
Однако в то же время все больше и больше генов отключаются из-за метилирования, что приводит к дальнейшей специализации клеток внутри клеточной линии. В процессах репарации ферменты репарации основаны на исходной цепи ДНК, которую они распознают благодаря более высокой степени метилирования. Дополнительная цепь также строится на основе хранящейся там информации. Ошибки при установке исправляются сразу. Кроме того, фермент AID (индуцированная активацией цитидин дезаминаза) очень специфично катализирует дезаминирование цитидиновых групп до уридиновых групп в одноцепочечной ДНК. Происходят соматические гипермутации, которые изменяют последовательности антител В-клеток. Затем выбираются подходящие B-клетки. Это дает возможность гибкого иммунного ответа.
Образование, возникновение, свойства и оптимальные значения
Цитидин является промежуточным продуктом метаболизма пиримидина. В качестве изолированного соединения это не имеет значения. Как уже упоминалось, он состоит из нуклеинового основания цитозина и пятикратной сахарной рибозы. Организм может сам синтезировать цитозин.
Однако его синтез очень энергоемкий, так что он восстанавливается из строительных блоков нуклеиновых кислот в рамках пути спасения и может быть реинтегрирован в нуклеиновые кислоты. Когда основание полностью разрушается, образуется диоксид углерода, вода и мочевина. Он присутствует в РНК как нуклеозид. В ДНК цитозин связан с дезоксирибозой, поэтому нуклеозид дезоксицитидин выступает здесь в качестве строительного блока.
Заболевания и расстройства
Метилирование цитидиновых остатков ДНК очень важно для маркировки, чтобы разделить различные биохимические процессы. Однако при метилировании также могут возникать ошибки, приводящие к заболеваниям.
В случае дефектного метилирования могут быть вызваны как повышенная, так и пониженная активность генов, не отвечающая требованиям. Эти паттерны метилирования передаются во время деления клеток. В долгосрочной перспективе происходят изменения, которые могут привести к заболеваниям. Например, некоторые опухолевые клетки имеют разные структуры метилирования, которых нет в здоровых клетках. Например, метилирование может блокировать определенные гены, кодирующие ферменты, регулирующие рост. Если эти ферменты отсутствуют, может произойти неограниченный рост клеток. Это также относится к ферментам, которые инициируют упорядоченную гибель клеток (апоптоз) при возникновении дефектов клеток.
На сегодняшний день целенаправленное влияние на метилирование ДНК пока невозможно. Тем не менее, существуют исследования полного деметилирования опухолевых клеток, чтобы снова подвергнуть их контролю белков, регулирующих рост. Согласно нескольким клиническим исследованиям, деметилирование ограничивает рост опухоли у пациентов с острым миелоидным лейкозом. Эта процедура также известна как эпигенетическая терапия. Процессы метилирования также могут играть роль при других заболеваниях. Под влиянием окружающей среды организм адаптируется к изменившимся условиям с образованием биологических модификаций, основанных на метилировании остатков цитидина в ДНК. Таким образом, организм осуществляет процесс обучения, который, однако, также может вызвать неправильное регулирование.



.jpg)



.jpg)




.jpg)
.jpg)